|
|
|
Copyright ©1998 by The Resilience Alliance*
Adina Merenlender, Claire Kremen, Marius Rakotondratsima, and Andrew Weiss. 1998. Monitoring Impacts of Natural Resource Extraction on Lemurs of the Masoala Peninsula, Madagascar. Conservation Ecology [online] 2(2): 5. Available from the Internet. URL: http://www.consecol.org/vol2/iss2/art5/
A version of this article in which text, figures, tables, and appendices are separate files may be found by following this link.
Research Monitoring Impacts of Natural Resource Extraction on Lemurs of the Masoala Peninsula, Madagascar Adina Merenlender1, Claire Kremen2, Marius Rakotondratsima3, and Andrew Weiss4
1University of California, Berkeley; 2Center for Conservation Biology and Wildlife Conservation Society; 3Wildlife Conservation Society, Madagascar; 4Center for Conservation Biology
- Abstract
- Introduction
- Background Information and Study Sites
- Methods
- Census Methods
- Demography
- Density, Physiography, and Population Size
- Results
- Census Methods
- Sample Effort
- Demography
- Density and Population Size Estimates
- Discussion
- Demography and the Influence of Resource Extraction
- Density and Population Size Estimates
- Novel Methods
- Responses to this Article
- Acknowledgments
- Literature Cited
Monitoring the influence of human actions on flagship species is an important part of conserving biodiversity, because the information gained is crucial for the development and adaptation of conservation management plans. On the Masoala Peninsula in Madagascar, we are monitoring the two largest prosimian species, Eulemur fulvus albifrons and Varecia variegata rubra, at disturbed and undisturbed forest sites to determine if extraction of forest resources has a significant impact on the population viability of these species. To test the sensitivity of lemur species to routine extraction of natural resources by local villagers, we compared population demography and density for both species across six study sites, using a new census technique. Three of the study sites were closer to villages and, therefore, were more impacted by resource extraction than the others. Our data on more than 600 individual primates suggest that the level of resource extraction did not significantly influence group size, fecundity, or density for either species over the two-year period of this study; however sex ratios in Eulemur were biased toward juvenile and adult females in more disturbed areas, suggesting that males may be emigrating from areas of less suitable habitat. Population densities at each site and estimates of population size across the entire peninsula were calculated and used to evaluate the design of a new park in the area, and to ensure that it will be large enough to support viable populations of these threatened primates. These estimates were calculated by obtaining the surface area of each study region from a geographic information system. Monitoring of these species continues in buffer zone areas of the park, where resource extraction is still permitted.
KEY WORDS: biodiversity, buffer zone, census techniques, conservation, Eulemur fulvus albifrons, GIS, lemurs, Madagascar, national park, natural resource extraction, primate, protected area management, Varecia variegata rubra.
The forests of Madagascar are relatively depauperate of large vertebrates compared to continental tropical forests. Thus, arboreal lemurs of 2.5 kg or more are among the largest vertebrates on the island, and have relatively large territories (Rigamonti 1993, Sterling 1993, Vasey 1996). Ensuring their survival may benefit other animals and plants with more restricted ranges. Many lemurs are frugivorous and serve as important seed dispersers for forest trees (White 1989, Rigamonti 1993) and pollinators of some forest canopy flowers (Kress et al. 1994). Lemurs may also be a food source for birds of prey and terrestrial carnivores (Langrand 1990, Mittermeier et al. 1994). Because of the ecological roles that they play, a reduction in lemur distribution and abundance could alter the structure, composition, and function of the forest community. Because lemurs are flagship species in Madagascar, they are an appropriate taxon to help monitor the success of current efforts in Madagascar to protect intact, functioning ecosystems and to manage forest resources sustainably (Kremen et al. 1994).
Lemurs currently suffer from three major threats in Madagascar. Conversion of forests for agriculture has already led to extreme habitat loss and fragmentation. Only about 10% of the native vegetation of Madagascar remains (Nelson and Horning 1993, Du Puy and Moat 1996), and rain forest conversion continues at an estimated 1.5% per year (Green and Sussman 1990). Low-level habitat alteration that results from selective extraction of timber and non-timber forest products may also influence lemur population viability. Selective extraction for subsistence or commercial use alters both forest structure and composition, potentially influencing the ability of large lemurs to move through the forest, and removing nest sites and food trees (Johns 1992, White et al. 1995). Finally, many lemur species are also hunted for food (Nicoll and Langrand 1989, Mittermeier et al.1992), which, if severe, can have consequences for population viability.
In this paper, we focus on the impact of forest product extraction on populations of the two largest lemurs of the Masoala Peninsula in northeastern Madagascar (Fig. 1) as an aid in designing a new national park and forest management zone for the region (Kremen et al. 1998; Kremen et al. in review). Although the creation of this national park will reduce threats to lemurs from habitat destruction and hunting, the impact of forest product extraction on lemurs will continue, especially within the forest management zone around the park. Over the long term, changes in lemur densities due to selective natural resource extraction may negatively influence efforts to manage these forests sustainably for production of subsistence and commercial forest products, because lemurs are important pollinator and dispersal agents. Monitoring the response of lemurs to extraction therefore becomes a critical element not only for lemur conservation, but also for the forest management plan, providing information to adaptively alter the plan in years to come (Walters and Holling 1990).
Fig. 1. Photograph of rain forest on the Masoala Peninsula, Madagascar.

|
Simply comparing the abundance of lemurs at different locations is unlikely to allow us to detect the impacts of forest product extraction on population viability. For example, more disturbed (e.g., more heavily extracted) forests may provide enough structure and food for lemurs to exist, but not to sustain a positive growth rate. Such areas, referred to as population sinks, occur when a species inhabits subquality habitat only by virtue of continual immigration from an adjacent source population found in more suitable habitat (Pulliam 1988, Dias 1996). If the area of forest used by human inhabitants is adjacent to a large region of undisturbed interior forest, this utilized area might be a sink, continuously drawing animals away from the undisturbed source area. Densities might, therefore, be similar between the two areas, without reflecting the underlying unsuitability of the disturbed habitat. This can present a problem for population viability if source habitat continues to be converted to sink habitat, such as could be the case if natural resource extraction were to expand beyond protected-area buffer zones.
Other population measures must be studied in order to observe the influence of subquality habitat on lemur populations. For example, Lemur fulvus spp.have a social system that includes female philopatry and male dispersal (Merenlender 1993). Therefore, we may expect group size to be smaller and female-biased in areas where resources are limiting, because primate group size is often correlated with food patch size and abundance; limitations in food may reduce immigration by males (Terborgh 1983, Ganzhorn 1988). Population sinks are best identified by long-term demographic studies rather than by densities (Pulliam 1988).
The hypothesis that disturbed forests represent population sinks for lemurs was tested by comparing demographic parameters for the two largest lemur species found on the Masoala Peninsula, Varecia variegata rubra (Fig. 2) and Eulemur fulvus albifrons (Fig. 3), in three sites located near human settlements and three sites farther away. We report our measurements of group size, composition, and reproductive success of all social groups at these sites for the period 1993 -1995, as well as estimates of density and population size. This monitoring program continues annually to allow observation of long-term trends in population dynamics.
|
Fig. 2. Photograph of Varecia variegata rubra by David Haring.

|
|
Fig. 3. Photograph of Eulemur fulvus albifrons by
David Haring.

|
Subspecies of Eulemur fulvus are abundant in forests across Madagascar (Tattersall 1982) whereas V. v. rubra is endemic to the Masoala Peninsula and is found at relatively low densities (Lindsay and Simons 1986, Mittermeier et al. 1992, White et al. 1995). Varecia v. rubra and E. f. albifrons may respond differentially to changes in forest composition resulting from human resource extraction because of their differences in size and diet. Varecia v. rubra is chiefly a frugivore, depending primarily on only a few fruiting trees (Rigamonti 1993), whereas E. fulvus is more of a generalist, eating a wide variety of fruits, flowers, insects, and leaves (Overdorff 1991). Both lemur species consume many plants that are also harvested by humans. We estimate that the overlap between plant species used both by lemurs and humans is approximately 50%, based on studies of lemurs' diets (Morland 1991, Rigamonti 1993; A. Merenlender, C. Kremen, M. Rakotondratsima, and A. Weiss,unpublished data) and information on resource extraction by humans on Masoala (Raymond 1995). The two species also have very different life history strategies, which will influence how they respond to habitat modification (Tattersall 1982). For example, Varecia have multiple offspring and use nests, whereas Eulemur usually have one offspring at a time and carry the infant (Morland 1992, Overdorff 1996). Therefore, using both species as indicators and comparing them will lead to a better understanding of the influence of forest product extraction on forest ecology.
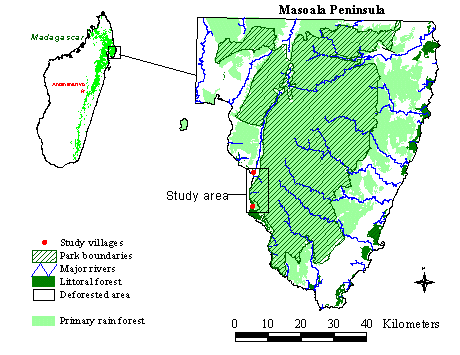
Background Information and Study SitesMadagascar is among 12 top-priority countries for biodiversity conservation (Myers 1988), supporting a highly unique flora and fauna with species-level endemism exceeding 75% in many taxonomic groups (Nicoll and Langrand 1989, Glaw and Vences 1994, Mittermeier et al. 1994, Dransfield and Beentje 1995). Recognizing the national and global importance of its biodiversity, the Malagasy government has embarked on an ambitious conservation program that proposes to establish 14 new protected areas (World Bank 1988). The Masoala Peninsula, a 4625- km2 area, covered approximately 75% in primary rain forest, was slated as one of these regions because it contains the largest remaining forested area in Madagascar. A park of 2100 km2 was proposed to the Madagascar government in 1995, and was signed into law in 1997 (Fig. 4). The park is part of an integrated conservation and development project (Wells et al. 1992, Wells and Brandon 1993, Kremen et al. 1994); this project is working with local people to protect the 1000 km2 of forests outside of the reserve as an extractive reserve for sustainable natural forest management.
Fig. 4. Map of the Masoala Peninsula, Madagascar, and location of study area.
|
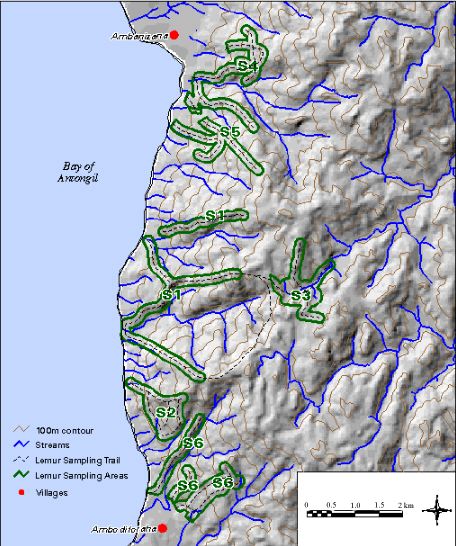
Study sites were located on the west coast of the Masoala Peninsula between the village of Ambanizana and the Tampolo Point (Fig. 4). This area spans 13 km of coast line, with a rugged, west-facing topography rising steeply from sea level to 700 m elevation within 3 km of the coast (Fig. 5). We selected six forested areas, half being far from villages and, thus, receiving limited or no human visitation, and half adjacent to villages and receiving regular visitation from local people, who harvest a wide variety of forest products (Raymond 1995). In addition to distance from the village, the existence of trails, which signify regular human activity in the area, was another criterion for distinguishing between impacted and unimpacted forested areas. The undisturbed sites had very limited or no preexisting trail system, whereas those classified as disturbed had well-used trail systems. We are in the process of quantifying the amount and type of resources removed from the forest by local villagers at various distances along trails, close and far from villages. We will then be able to compare these data with information from village surveys on natural resources that are regularly extracted to compile a complete picture of forest product extraction. Although the length of trail used for censusing and the areas were different at each site, the total amount of trails (16.7 km and 16.8 km) and area (1462 ha and 1312 ha) for undisturbed and disturbed forest sites were almost identical.
Fig. 5. Map of each study site.
|
Although local villagers hunt lemurs, especially in areas boarding agricultural fields and forest, such hunting was uncommon at our study sites. Therefore, the direct removal of animals by humans was not a frequent event and, therefore, most likely is not the cause of differences found in primate demography or density between localities. In the future, continued presence of researchers, the integrated conservation and development project, and the establishment of a national park will help to deter these illegal practices.
All sites were located in lowland humid evergreen forest, typified by Canarium (Burseraceae), Diospyros (Ebenaceae), Symphonia (Clusiaceae), Tambourissa (Monimiaceae), Weinmannia (Cunoniaceae), and Anthostema and Uapaca (Euphorbiaceae), among others (Rakotondratsima 1995, Kremen et al. 1998). Table 1 presents physical characteristics of each site. Five out of the six research sites had very similar elevational range and aspect. Site S3 was the only study site that is east of the north - south escarpment and is located in the interior forest of the peninsula. This resulted in higher mean elevation and a decrease in elevational range at the site, as well as a different average aspect (Table 1).
Table 1. Physiographic characteristics of all research sites.
a. Distance from the nearest village.
|
The primate census techniques that we used differ in several ways from classic methods (Southwick and Cadigan 1972, Struhsaker 1975, Defler and Pintor 1985) and allow low-cost, long-term monitoring. Our method relies on (1) repeatedly identifying social groups, and (2) obtaining demographic data on all identified groups. The method thereby permits us to obtain a large quantity of demographic data reliably for many groups across many sites, and is therefore an appropriate method for monitoring changes in primate abundance and demography.
In 1993, sites were selected, trails were marked at 50-m intervals, two local villagers were trained in collecting data, and lemur groups of E. f. albifrons and V. v. rubra were identified at each site. However, because the 1993 data were collected during the training and initial census period, and two additional sites were added at the end of the year, we present the 1993 data for the analysis of sample effort only. Demographic and density data are presented for 1994 - 1995, once all groups were already identified and field staff were fully trained.
At most sites, sampling was conducted on pre-existing trails that had either been created by villagers for forest product extraction, or by researchers working in the area. Most pre-existing trails were found along ridge lines. Using ridge line trails while censusing lemur groups is advantageous because calls can be heard well from the ridge tops and visibility of the slope below is often better. Some trails did run perpendicular to the ridge line in order to better penetrate the forest covered slopes (Fig. 5). Use of pre-existing trails is also desirable in this region, because cutting of new trails can stimulate additional forest product extraction in previously untouched areas.
Each trail was followed from early morning to early afternoon, at a slow pace, by two people looking for animals. During the census period, field-workers tried to find all detectable groups and to count, sex, and determine the age class whenever possible. Once a group of similar composition, or with identifiable individuals, was seen at approximately the same place three times, it was given an identification number; subsequent sightings of the group were listed by this group number. The data collected at each sighting were: (1) group number, (2) total group size seen, (3) number of adult, juvenile, and infant females and males, (4) closest trail marker, (5) approximate distance from the trail marker to where the animals were found, and (6) notes that included behavioral observations and feeding records. These data were taken over a three-day period from five to eight times at each site in 1994. From the work accomplished in 1994, it was determined that the local field technicians taking the data were extremely familiar with the groups being monitored, and continued censuses could be restricted to the months following the birth season, without loss of accuracy. In 1995, therefore, the sites were visited from two to six times only, depending on the difficulty involved in getting to the site.
These census methods differ from the traditional line-transect sampling methods (Whitesides et al. 1988), in that the observers regularly left the trail to identify lemur groups up to 100 m from the trail, to obtain the best possible data on group size and composition. A comparison of our methods with the standard line-transect sampling was made during the middle of the field season (December1995 - January 1996) at two disturbed (S4 and S6) and two undisturbed (S2 and S3) sites. This was done to assess the quality and accuracy of data calculated using our field methods as compared to more standard primate censusing techniques. Data for the line-transect method were taken while remaining on the trail (transect), and the following calculations were compared between the two census techniques: (1) number of individuals observed, (2) total group size, (3) sex ratio, and (4) density.
DemographyThe primary demographic parameters estimated from the repeated census data were group size, sex ratio, and female fecundity. To determine the group size and composition of the identified E. f. albifrons groups, the maximum group size recorded during the field season and group composition at that time were used in the comparisons for each year. This was done to avoid using incomplete group counts that can result from poor weather or dense canopy. Sex of the individuals censused was recorded for E. f. albifrons. For V. v. rubra, each subgroup whose members were regularly seen together was identified as a separate group. We did observe that neighboring subgroups sometimes formed larger social groups, and this has been observed in Varecia by other workers (Morland 1991, Rigamonti 1993). Most of the visits to our study sites were made during the dry season, because it is easier to observe lemurs at that time. During this season, V. v. rubra were often observed in small groups of 2 - 4 animals. We hypothesized that these were subsets of larger, cohesive social groups, and thus, we could not determine social group size for V. v. rubra. We also could not determine sex ratio for this species, because distinguishing the sex of individual V. v. rubra in the field is unreliable using rapid census methods.
Average female fecundity for E. f. albifrons was calculated, for each social group, as the number of infants born divided by the number of females thought to be more than two years old. Because we could not reliably sex adult V. v. rubra, we assumed an even adult sex ratio, as has been observed in other Varecia populations (Morland 1992). Reproductive success was then calculated for each group as the total number of infants born in the group, divided by half of the number of adults in the group. Varecia infants are born in the dry season around October, and remain in a nest for about 3 months following birth (Morland 1990). Varecia v. rubra infants were not generally observed during censusing before December; therefore, the 1993 infants were first observed early in 1994, and the 1994 infants were first observed in early 1995.
Nonparametric analysis of variance and paired tests were used to examine demographic parameters across all research sites. Although nested ANOVAs, with disturbance as level 1 and site as level 2, would have been the preferred method, significant heteroscedasticity was detected. Therefore, Kruskal-Wallis ANOVA and the Wilcoxon matched-pairs signed-rank test are reported instead of parametric tests. This choice did not affect the conclusions.
Density, Physiography, and Population SizeAnimal density was calculated by dividing the number of adult and juvenile animals in all of the identified social groups at each site by the study site area. The study site areas were estimated using GIS coverages of lemur census trails and a Digital Elevation Model (DEM) of the study area. The DEM, with a 30-m horizontal resolution, was developed using the Arc/Info 7.1.1 TOPOGRID command (ESRI 1996, Hutchinson 1996), with inputs of 25-m contours, spot elevations, coastlines, and hydrology digitized from Malagasy 1:100,000 scale maps. Trails were mapped using a global positioning system (GPS), and were improved with geographic references to topography, hydrology, compass, and altimeter readings (Fig. 4). This additional information was needed to correct GPS readings, because no base station data are available for Masoala, preventing differential correction of GPS data. These trails were buffered at a given distance using the Arc/Info 7.1.1 GRID command PATHDISTANCE (ESRI 1996), which computes for each location the minimum distance from a trail, taking into account slope distance. Surface area within the buffers was calculated by dividing the planimetric area of each cell (900 m2) by the cosine of the slope.
We determined the distance to use for buffering the sampling trails by using estimated home ranges documented in similar habitat. Eulemur fulvus rufus in eastern Madagascan rain forest has a core home range of 40 ha (non-overlapping territories) within a total range of 100 ha (overlapping territories) (Overdorff 1991, 1996). If core and total home ranges are modeled as circular areas, they then have radii of 356 - 564 m. For this reason, we buffered trails at distances of 350m and 500 m for E.f. albifrons, and estimated an upper and lower density based on the surface area included within these buffers. Similarly, we used published estimates of home range size for Varecia from two studies (Rigamonti 1993, Vasey 1996) on the Masoala Peninsula. Home range estimates in the two studies varied from 24 ha to 57.7 ha, or circular radii of 89 - 428 m. We therefore used trail buffers of 100 m and 500 m to calculate upper and lower bound density estimates for Varecia. The physiographic characteristics of study sites, such as elevation and aspect, were then calculated from the topography coverage using the largest buffer distance (500 m).
To calculate the most conservative estimate of total population size for the two lemur species on the entire Masoala Peninsula and within the future park, the density estimates based on 500-m buffers were multiplied by estimates of the area of forest found on the peninsula or within the park. The entire area of primary forest was used for these calculations, because both species have been observed in all primary forest types known to occur on Masoala, and are present ubiquitously across the peninsula (Sterling and Rakotoarison 1998). The forested area of the Masoala Peninsula was calculated from a SPOT panchromatic image (10-m resolution) from 1991, which readily distinguishes deforested from primary forest areas. Surface area of primary forest was then calculated for the entire peninsula and for the core park area, based on a Digital Elevation Model from 100-m contours.
Our census methods demonstrated a higher detection rate of individual lemurs than the standard line-transect methods, which is an advantage for collecting demographic data. There are also differences in density estimates for E. f. albifrons as a result of the different techniques used. The total number of individuals observed over the 19-day census period at four study sites across 38 km of trail (transects) for each species was significantly larger using our census methods (t = 3.05, df = 18, P < 0.01) (E. f. albifrons: our methods N = 240, line-transect N = 186; V. v. rubra: our methods N = 84, line-transect N = 75). Group size was also significantly larger for E. f. albifrons using our method (t = 3.98, df = 28, P<0.01) (mean = 8.28, our method; mean = 6.41, line-transect method). Line-transect methods are designed to estimate density using an estimated census drop-off distance, based on the period of observation and length of the transect. Using the line-transect method with a drop-off distance of 45.8 m, the population density for E. f. albifrons is 106 individuals/km2; for V. v. rubra, it is 7.12 individuals/km2, using a drop-off distance of 300 m.
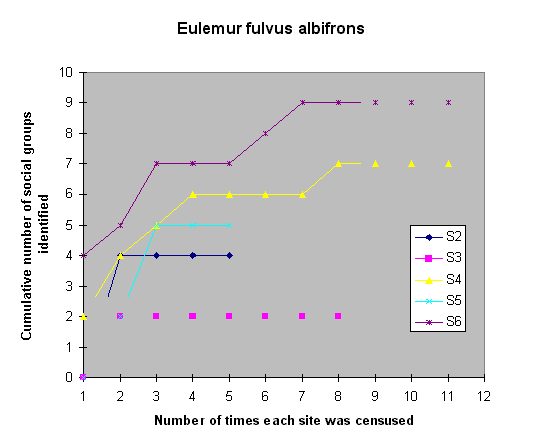
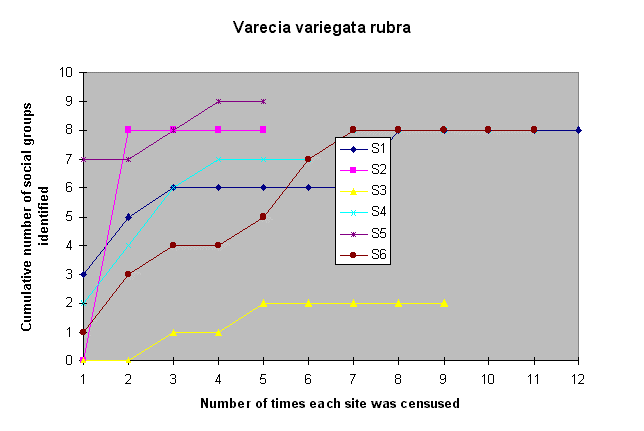
Sample EffortFigures 6 and 7 show the number of visits required to identify the lemur groups, found at each site during the initial 1993 census, that were repeatedly censused throughout the study. Field staff identified 77 - 100% of the social groups ofE. f. albifrons by the fifth visit (Fig. 6), and 62 - 100% of the social groups of V. v. rubra (Fig. 7). For either species, eight visits were required at one of the six sites to identify all of the social groups.
Fig. 6. Sample effort curve for Eulemur fulvus albifrons, demonstrating the number of social groups that were identified after consecutive visits to the study sites.
|
Fig. 7. Sample effort curve for Varecia r. rubra,
demonstrating the number of social groups that were
identified after consecutive visits to the study sites.
|
Demography
We identified approximately 428 individual E. f. albifrons in 1994 and 448 in 1995, in 45 individual social groups across six study sites (Table 2). We identified 48 V. v. rubra groups with a total of 185 individuals in 1994, and 188 in 1995 (Table 2). Kruskal-Wallis ANOVA revealed no overall significant effect of disturbance (1994: H(1, N =45) = 0.56, P = 0.45; 1995: H(1, N = 45) = 0.18, P = 0.67) or study site on E. f. albifrons group size (1994: H(5, N = 45) = 2.67, P = 0.75; 1995: H(5, N = 45) = 2.72, P = 0.74). Wilcoxon matched-pairs tests did not reveal any significant changes in overall group size from 1994 to 1995, at all six study sites, for both species.
Table 2. Total number of lemur groups censused, total number of animals censused, and group size (mean + 1 SE) for each of six sites in 1994 and 1995.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
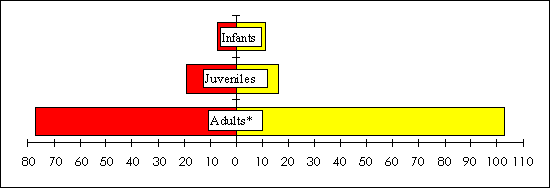
The following results are for E. f. albifrons, the sexually dichromatic species. The only site that demonstrated a significant female bias for juveniles in 1994 (chi-square = 3.5, df = 1, P<0.06) and adults in 1995 (chi-square = 5.5, df = 1, P< 0.02) was S5, a site with high levels of natural resource extraction. Further, when all disturbed sites were lumped together, there was a significant female bias for juveniles in 1994 (chi-square = 30, df = 1, P<0.001; Fig. 8) and adults in 1995 (chi-square = 3.75, df=1, P<0.05; Fig. 9). In the less disturbed sites, there was no significant difference from an even sex ratio.
Fig. 8. Total number of male (left axis) and female (right axis) Eulemur f. albifrons found in the three less disturbed study sites in 1994. The asterisk indicates a significant deviation from a 1:1 sex ratio at P < 0.001.
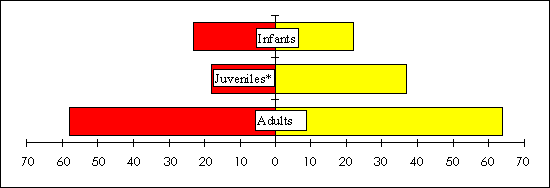
|
Fig. 9. Total number of male (left axis) and female (right
axis) Eulemur f. albifrons found in the three
more disturbed study sites in 1995. The asterisk indicates a
significant deviation from a 1:1 sex ratio at
P < 0.05.
|
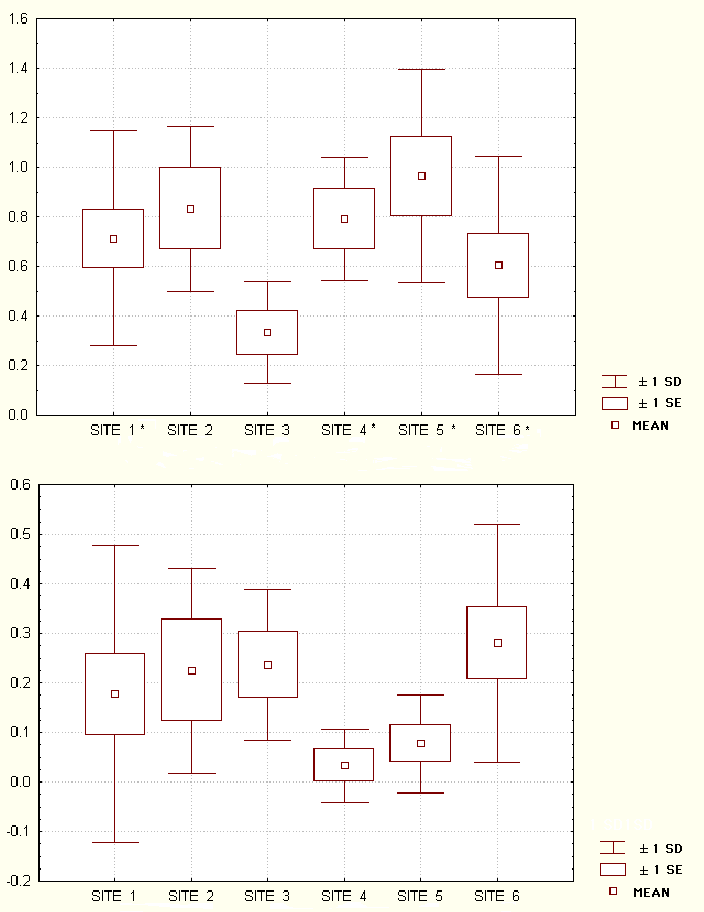
The mean female fecundity for E. f. albifrons was calculated for each study site (Fig. 10). Kruskal Wallis ANOVA revealed no overall significant effect of disturbance (1994: H(1, N = 45) = 0.15, P = 0.70; 1995: H(1, N = 45) = 0.19, P = 0.66) or study site on E. f. albifrons fecundity (1994: H(5, N = 45) = 7.98, P = 0.15; 1995: H(5, N = 45) = 8.72, P = 0.12). The same was true for V. v. rubra for the effect of disturbance (1994: H(1, N = 49) = 0.30, P = 0.58; 1995: H(1, N = 49) = 0.00, P = 0.97) and study site on fecundity (1994: H(1, N = 49) = 0.1.2, P = 0.94; 1995: H(1, N = 49) = 3.4, P = 0.63). However, in both years, the social groups at S3 had a lower mean fecundity rate than that found at the other sites. There was a significant decrease in female fecundity from 1994 to 1995 at most of the study sites, as shown by the Wilcoxon rank sum for the Wilcoxon's matched-pairs signed-ranks test (Fig. 10).
Fig. 10. Female fecundity values for Eulemur f. albifrons at each study site. Fecundity is measured as the number of infants born divided by the number of females thought to be greater than two years old in 1994 and 1995. Asterisks indicate significant results (P<0.07) from Wilcoxon matched pairs between years.
|
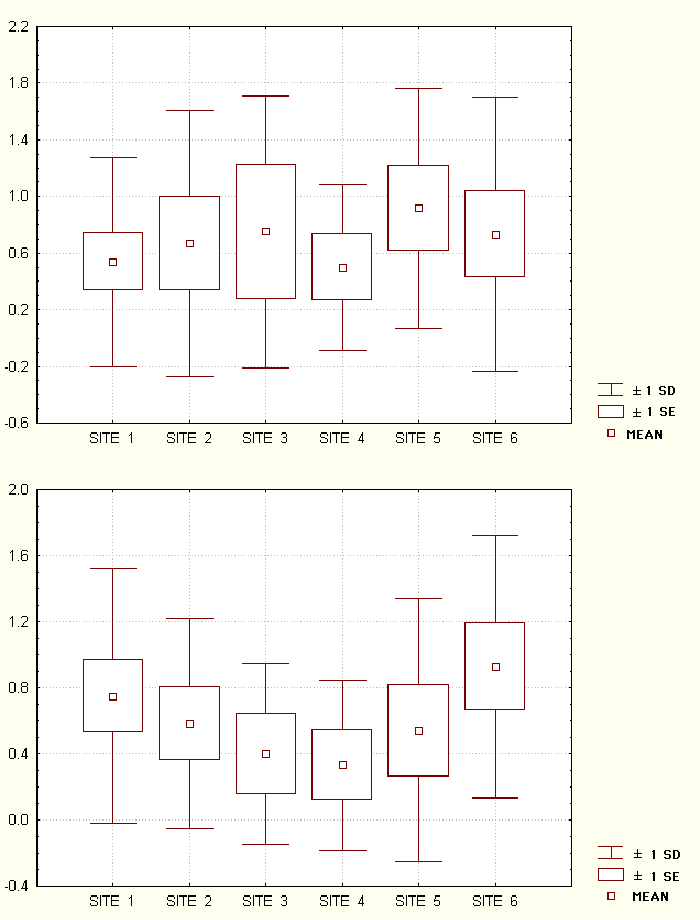
Reproductive rate for V. v. rubra was not influenced by study site in 1994 (df = 5, N = 49, H = 1.22, P = 0.94) or 1995 (df = 5, N = 48, H = 3.43, P = 0.63). There is no significant trend of either increasing or decreasing fecundity between 1993 and 1994 for V. v. rubra at all of the study sites (Fig. 11). However, a single group rarely reproduces in consecutive years. Out of 48 groups censused at all six sites for the two years, we recorded 17 groups only having infants in 1993, 20 groups only having infants in 1994, 4 groups having infants in both years, and 7 groups not having infants in either year.
Fig. 11. Female fecundity values for Varecia v. rubra at each study site. Fecundity is measured as the number of infants born divided by the number of females thought to be greater than two years old in 1994 and 1995.
|
Density and Population Size Estimates
Lower and upper mean densities of E. f. albifrons for the six sites, based on the two trail buffers (see Methods), ranged from 15.67 to 21.52 animals/km2 in 1994, and 16.56 to 22.73 animals/km2 in 1995 (Table 3). For V. v. rubra, densities ranged from 7.36 to 27.28 animals/km2 in 1994, and 7.56 to 28.02 animals/km2 in 1995.
Table 3. Minimum and maximum densities (no. animals/km2 ) of E. f. albifrons and V. v. rubra found on the Masoala Peninsula at the six study sites in 1994 and 1995, as estimated using areas calculated from a variable buffer width around sampling trails.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
No significant differences were found between the disturbed and undisturbed sites in densities of E. f. albifrons or V. v. rubra during either 1994 or 1995, for either the minimum or maximum estimates. Paired t tests between 1994 and 1995 reveal that the density estimates between years are significantly different (E. f. albifrons, 350-m buffer: ts = 2.9 , df = 5, P < 0.05; E. f. albifrons, 500-m buffer: ts = 2.9 , df = 5, P < 0.05; V. v. rubra, 100-m buffer: ts = 3.7 , df = 5, P < 0.05; V. v. rubra, 500-m buffer: ts = 4.29, df = 5, P<0.05). For E. f. albifrons, density increased from 1994 to 1995 across all sites, whereas V. v. rubra did not display the same trend across all sites.
The most conservative 1995 mean ( + 1 SE) density estimates, based on 500-m buffers, result in estimated population sizes for V. v. rubra of 16,179 + 3254 and 35,451 + 4715 for E. f. albifrons within the park alone, and 25,542 + 5137 and 55,967 + 7444 for the entire forested area, respectively.
Eulemur is one of the most widespread prosimian genera in Madagascar; its members are presumed to be resilient to moderate levels of forest disturbance (Tattersall 1982). Our results for E. f. albifrons support this, because we were able to detect only a slight difference in group size and composition between forested areas neighboring villages and undisturbed sites on the west coast of Masoala. In contrast, others have suggested that Varecia are sensitive to forest health (White et al. 1995) and, therefore, may suffer from natural resource extraction. This notion was not supported in our study of V. v. rubra, because we were not able to differentiate female fecundity and density between disturbed and undisturbed forested areas from the available data, although we did not include dramatically altered forests in our monitoring program. There are forest patches in Madagascar that have been impacted to the point at which few or no lemurs are supported. Additional years of data may be required in order to detect a significant trend for reproductive success in protected versus utilized forest patches. However, our data suggest that low levels of forest resource extraction such as those found on Masoala may pose little danger to lemur populations.
These preliminary results are encouraging, in that the regions of forest left outside of the park (123,865 ha) will probably continue to provide habitat for lemur populations, as long as forest cover is maintained and the ban on hunting lemurs is enforced. These areas were purposefully left outside of the park as a buffer to the park; in this zone, marketing of timber and non-timber forest products is being promoted as a favorable alternative to agricultural expansion (Kremen et al., in review). Without some measure to increase the value of standing forest to local people and to provide for them economically , agricultural expansion would, in time, devastate not only these border forests, but also the forests within the park, as has happened elsewhere in Madagascar (Smith et al.1997). The use of these border forests for intensified extraction may ultimately lower their habitat suitability for lemurs and other creatures, but it provides a key element in the overall conservation and resource management strategy for the entire peninsula. Monitoring of these two lemur species at these and additional study sites is continuing in order to assess the effectiveness both of the conservation program and the impacts of intensified extraction in the border forests.
Demography and the Influence of Resource ExtractionReliable demographic data are required for primate species across space and time, in order to assess the value of protected areas in conserving primates and to monitor the impacts on primates of habitat management activities, including resource extraction in unprotected areas or extractive reserves. Demographic variables of primates are often highly variable across sites, habitat types, social groups, and individuals (Altmann et al. 1985, Richard et al. 1991, Sussman 1991); yet, many primate studies have focused on only one or a few groups, so statistically reliable estimates of demographic parameters are infrequent in the literature (Dobson and Lyles 1989). This is unfortunate, because data on population demography are more likely to reveal the effects of habitat management activities on primate populations.
Primate group size is often associated with resource distribution and abundance (Terborgh 1983, Ganzhorn 1988, Chapman 1990, Janson 1992). Primate groups found in disturbed or marginal forests may be smaller when available resources are limited, or larger if regenerating forests produce increased food resources (Ganzhorn 1988, Olupot et al. 1994). However, analysis of our preliminary results did not detect a significant difference in group size between study sites that are differentially affected by natural resource extraction by villagers.
Female fecundity can vary among individuals, social groups, years, and habitat. Little information exists on fecundity rates of any lemur species for many individuals across a large landscape (Sussman 1991). Our study, therefore, represents one of the most comprehensive monitoring efforts of lemur fecundity in the wild. In this study, we did not detect an effect of human disturbance on fecundity in either species. This suggests that individual and social group differences explain more of the variation in fecundity rates than do the differences in habitat integrity or forest type differences that were the focus of this study. In E. f. albifrons, we found a significant between-years difference in female fecundity at most sites. This supports an earlier study that showed a difference in birth rate between years for individual female Eulemur elsewhere in Madagascar (Overdorff et al., in review). Similarly, our findings support an earlier suggestion that Varecia successfully reproduces only once in two years in the wild (Morland 1990). This birth interval may be due to the fact that Varecia regularly give birth to several infants at once and often successfully raise more than one in the wild, which requires a greater expenditure of energy than for those prosimians who raise a single infant with an interbirth interval of one year (Tattersall 1982).
Available food resources and forest structure can impact demographic parameters, such as survivorship and dispersal patterns, differentially between males and females, thereby altering the sex ratio in primate populations. Although we did not have enough data to analyze infant sex ratio statistically, we know from captive studies and other field studies that E. fulvus has an even birth sex ratio (Watson et al. 1996, Overdorff 1991, Overdorff et al., in review). Differences in mortality and dispersal between the sexes influence the sex ratio of older age classes. Therefore, we believe that the female-biased sex ratio in the older age classes at the disturbed sites could be due either to greater resident male dispersal relative to male immigration, or to increased male mortality relative to females. In Eulemur, females are usually philopatric and males disperse; thus, we hypothesize that males would disperse from areas where resources are inadequate, resulting in the observed female-biased sex ratio if disturbed sites are resource limited. In contrast, there is no evidence to support differential mortality between males and females in disturbed sites.
Except for the trend toward female-biased sex ratios in disturbed sites, we cannot detect, in these early monitoring efforts, a significant difference in lemur demography between forested areas adjacent to rural villages and those more distant areas. We do not have a high probability of committing a Type II error: power estimates based on our 1994 data are high for both group size and fecundity data. The power to detect a 4% or greater increase or decrease in group size is greater than 0.97 for both lemur species, and the power to detect a 4% or greater change in female fecundity for both species is greater than 0.9. These power estimates were calculated with a Type I error rate of 0.05 using monitor.exe (Gibbs 1998). Sensitivity analysis demonstrated that the results of this power analysis were not sensitive to some of the estimated input values that we supplied (e.g., the number of times a primate group was observed per site visit). Other studies often accept power levels that only allow for a high probability of detecting a 20% increase or decrease (Zielinski and Stauffer 1996). Even the well-studied California Spotted Owl (Strix occidentalis) only produced power estimates of between 0.13 and 0.64 for detecting a 4% change in population size (Taylor and Gerrodette 1993).
Density and Population Size Estimates
We did not detect a significant effect of human resource utilization on lemur densities. However, these results were based on a short time period and the forested sites that were more disturbed are also contiguous to a vast amount of undisturbed forest. We suspect that this region provides significant source populations of both lemur species, making it difficult to detect any differences in overall density between more and less disturbed sites (Dias 1996).
The population density estimates that we calculated for E. f. albifrons with our methods are lower than those found using the strip-line census method on the Masoala Peninsula, as measured by us and by Sterling and Rakotoarison (1998), or for E. fulvus sp. elsewhere in Madagascar (Morland 1991, Sterling and Rakotoarison 1998, Overdorff et al., in review). For V. v. rubra, the upper mean density estimate (based on 100-m buffers) is nearly identical to that found by Sterling and Rakotoarison (1998) in eastern Masoala.
Our estimates of population size for both lemur species on the entire peninsula and for the national park were based on the assumption that all forested areas on the peninsula provide suitable habitat for these species. Both species have been seen by field-workers throughout the peninsula; however, these estimates could be improved by data from additional localities across the peninsula, to take into account varying levels of animal abundance due to differences in the habitat types.
The estimates of population size that we arrived at are in the same order of magnitude as total population sizes calculated by extrapolating directly from published home range sizes. Using upper and lower home range sizes of 100 ha and 40 ha for E. f. albifrons (Overdorff 1991), with mean group sizes of 9.39 animals per group, based on our 1995 data, E. f. albifrons would attain population sizes on the Masoala Peninsula of 31,740 - 79,345 animals if all forested areas were fully occupied by social groups. Our population size estimate (mean + 1 SE) for the entire peninsula was 55,967 + 7444 animals. Similarly, using upper and lower home range estimates with corresponding average group sizes for V. v. rubra from published studies (Rigamonti 1993, Vasey 1996), we estimate population size on the peninsula to be 75,725 - 144,020 animals. Our population size estimate for the entire peninsula was 25,542 + 5137 animals, a more conservative estimate. We used these population size estimates to assess the design of the proposed Masoala National Park, concluding that the proposed park of 214,145 ha of forest would be large enough to protect significant populations of each species.
Novel MethodsGiven that over 90% of the world's primates are endangered, developing efficient methods for monitoring primates is an essential step for primate conservation biology. To monitor lemur populations, we employed several novel field and analytical methods. A low-technology method of identifying lemur social groups for long-term monitoring was developed to maximize the collection of data relevant to primate demography. However because individual animals are not marked, a certain level of error may have been introduced if observers either lumped or split groups. In order to measure this error for quality assurance purposes, we would have to mark several primate groups to allow a comparison of the data from marked vs. unmarked animals. This requires extensive animal handling that could not be accomplished as part of this project. However, integrated conservation and development projects would benefit greatly from effective quality control programs. Such a program would provide estimates of the level of error in biological and socioeconomic monitoring that is inherent in the data collection performed by the various participants in the project. For example, a forest health monitoring program in the United States measured the agreement between experienced trainers and inexperienced technicians who collected the species richness and cover data, and found the technicians' data to be highly precise (Stapanian et al. 1997).
A team of local villagers who were trained and supervised by us collected all of the field data. These methods take full advantage of indigenous peoples' knowledge of the forest flora and fauna. In the United States, it is widely recognized that monitoring environmental quality should involve local citizenry, leading to an increase in volunteer monitoring programs (Kerr et al. 1994). Often, the job of monitoring vast amounts of natural resources, such as regional water quality, requires large numbers of data collectors who can only be obtained through a volunteer program (Heiskary et al. 1994). Not only can trained, local residents provide valuable data, as was demonstrated in our study, but also the act of collecting ecological data increases their understand of the surrounding ecosystem and the importance of sustainable land use practices. This is exceedingly valuable, because the health of our environment should not rest with scientists and natural resource managers alone.
Our methods require less field time than traditional techniques of primate behavioral observation, but are more intensive than rapid survey methods. Traditional field primatology has emphasized the importance of individual animal behavior and social group dynamics which requires studying known (often marked) individuals and, thus, greatly restricts the number of animals and study sites (Dobson and Lyles 1989). Rapid surveys of primate density (Southwick and Cadigan 1972, Struhsaker 1975, Defler and Pintor 1985, Buckland et al. 1993, Sterling and Rakotoarison, in press) are also extremely limiting, because primate densities are usually low and primates are often elusive, thereby limiting the number of possible observations in a short field visit. The census methods we describe maximize the ability to (1) study many social groups of primates at a large number of study sites, (2) repeatedly identify resident lemur groups, and (3) quantify social group composition in addition to density. These new methods were also shown to improve detection rates relative to traditional line-transect methods. More individuals were observed and larger group sizes were recorded using our census methods, leading to a larger amount of demographic data. Despite better detection rates, our density estimates are lower than those found by other investigators in Madagscar (Morland 1991, Sterling and Rakotoarison 1998, Overdorff et al. in review). This suggests that the line-transect estimates are unrealistically high, particularly for E. f. albifrons. Density estimates of V. v. rubra, using our methods, are similar to those calculated using line-transect methods and those calculated by other investigators (Sterling and Rakotoarison 1998). These discrepancies demonstrate the difficulty inherent in estimating forest primate population densities.
The repeated census technique described here is useful for monitoring primate populations over space and time to provide statistical estimates of demographic parameters and to estimate population sizes. The use of geographic information systems permitted the characterization of study sites with respect to disturbance, physiography, and area, and also allowed freedom from the constraints of measured line-transect sampling, which severely limits field observations of animals, such as lemurs, that are patchily distributed across large areas. The use of GIS also allowed extrapolation of estimates of population size to a larger area. These methods could be adapted for monitoring other territorial animals. The combination of these methods has allowed us to estimate population density and to track demographic trends for multiple social groups of two lemur species at several research sites, with limited support funds. Robust estimates of these parameters can then be used to compare alternative habitat management schemes, and therefore provide input to an adaptive management process (Walters and Holling 1990, Kremen et al. 1994).
Responses to this article are invited. If accepted for publication, your response will be hyperlinked to the article. To submit a comment, follow this link. To read comments already accepted, follow this link.
Acknowledgments
Altmann, J., G. Hausfater, and S. A. Altmann. 1985. Demography of Amboseli baboons. American Journal of Primatology 8: 113-125.
Chapman, C. A. 1990. Ecological correlates of group size for three species of neotropical primates. Folia Primatologica 55:1-9
Defler, T. R., and D. Pintor. 1985. Censusing primates by transect in a forest of known primate density. International Journal of Primatology 6:243-259.
Dias, P. C. 1996. Sources and sinks in population biology. Trends in Ecology and Evolutionary Biology 11(8):326-329.
Dobson, A. P., and A. M. Lyles. 1989. Population dynamics and conservation of primates. Conservation Biology 3: 362-380.
Dransfield, J., and H. Beentje. 1995. The palms of Madagascar. Royal Botanic Gardens, Kew and The International Palm Society, London, UK.
Du Puy, D. J., and J. Moat. 1996. A refined classification of the primary vegetation of Madagascar based on the underlying geology: using GIS to map its distribution and to assess its conservation status. Pages 205-218 in W. R. Lourenco, editor. Biogeographie de Madagascar. ORSTOM, Paris, France.
ESRI (Environmental Systems Research Institute). 1996. Arc/Info Version 7.1.1 On-line documentation. Redlands, California, USA.
Ganzhorn, J. U. 1988. Food partitioning among Malagasy primates. Oecologia 75:436-450.
Gibbs, J. 1998. Monitoring program: Monitor.exe
http://www.im.nbs.gov/powcase/powcase.html
Glaw, F., and M. Vences. 1994. A fieldguide to the amphibians and reptiles of Madagascar. Second edition. Privately published, Koln, Germany.
Green, G. M., and R. W. Sussman. 1990. Deforestation history of the Eastern rain forests of Madagascar from satellite images. Science 248:212-215.
Heiskary, S., J. Lindbloom, and C. B. Wilson. 1994. Detecting water quality trends with citizen volunteer data. Lake and Reservoir Mangement 9(1):4-9.
Hutchinson, M. F. 1996. A locally adaptive approach to the interpoplation of digital elevation models. In Proceedings, Third International Conference/Workshop on Integrating GIS and Environmental Modeling Santa Fe New Mexico, 21-26 Jan. 1996. Santa Barbara, California, USA.
Janson, C. H. 1992. Evolutionary ecology of primate social structure. Pages 95-130 in E. A. Smith and B. Winterhalder, editors. Evolutionary ecology and human behavior. Aldine de Gruyter, New York, New York, USA.
Johns, A. D. 1992. Species conservation in managed tropical forests. Pages 15-53 in T. C. Whitmore and J. A. Sayer, editors. Tropical deforestation and species extinction. Chapman and Hall, London, UK.
Kerr, M., E. Ely, Y. Lee, and A. Mayio. 1994. A profile of volunteer environmental monitoring: National survey results. Lake and Reservoir Management 9(1):1-4.
Kremen, C., A. M. Merenlender, and D. D. Murphy. 1994. Ecological monitoring: a vital need for integrated conservation and development programs in the tropics. Conservation Biology 8:388-397.
Kremen, C. K., I. Raymond, and K. Lance. 1998. An interdisciplinary tool for monitoring conservation impacts in Madagascar. Conservation Biology 12:549-563.
Kremen, C., V. Razafimahatratra, R. P. Guillery, J. Rakotomalala, A. Weiss, and J. Ratsitsompatrarivo. In review. On scientific grounds: designing the Masoala National Park in Madagascar.
Kress, W. J., G. E. Schatz, M. Andrianifahanana, and H. S. Morland. 1994. Pollination of Ravenala Madagascariensis (Strelitziaceae) by lemurs in Madagascar: evidence for an archaic coevolutionary system. American Journal of Botany 81:542-551.
Langrand, O. 1990. Guide to the birds of Madagascar. Yale University Press, New Haven, Connecticut, USA.
Lindsay, N. B. D., and H. J. Simons. 1986. Notes on Varecia in the northern limits of its range. Dodo 23:19-24.
Merenlender, A. M. 1993. The effects of sociality on the demogrpahy and genetic structure of Lemur fulvus rufus (polygamous) and Lemur rubriventer (monogamous) and the conservation implications. Dissertation. University of Rochester, Rochester, New York, USA.
Mittermeier, R. A., W. R. Konstant, M. E. Nicholl, and O. Langrand. 1992. Lemurs of Madagascar: an action plan for their conservation 1993-1999. IUCN, Gland, Switzerland.
Mittermeier, R. A., I. Tattersall, W. R. Konstant, D. M. Meyers, and R. B. Mast. 1994. Lemurs of Madagascar. Conservation International, Washington, D.C., USA.
Morland, H. S. 1990. Parental behavior and infant development in ruffed lemurs (Varecia variegata) in a northeast Madagascar rain forest. American Journal of Primatology 20:253-265.
_______ . 1991. Preliminary report on the social organization of ruffed lemurs (Varecia variegata variegata) in a northeast Madagascar rain forest. Folia Primatologica 56:157-161.
_______ . 1992. Social organization and ecology of black and white and white ruffed lemurs in lowland rain forest, Nosy Mangabe, Madagascar. Dissertation. Yale University, New Haven, Connecticut, USA.
Myers, N. 1988. Threatened biotas: "hotspots" in tropical forests. The Environmentalist 8:1-20.
Nelson, R., and N. Horning. 1993. AVHRR-LAC estimates of forest area in Madagascar, 1990. International Journal of Remote Sensing 14:1463-1475.
Nicoll, M. E., and O. Langrand. 1989. Madagascar: revue de la conservation et des aires protégées. World Wide Fund for Nature, Gland, Switzerland.
Olupot, W., C. A. Chapman, C. H. Brown, and P. M. Waser. 1994. Mangabey (Cercocebus albigena) population density, group size, and ranging: a twenty-year comparison. American Journal of Primatology 32:197-205.
Overdorff, D. J. 1991. Ecological correlates to social structure in two prosimian primates: Eulemur fulvus rufus and Eulemur rubriventer in Madagascar. Dissertation, Duke University, Durham, North Carolina, USA.
_______ . 1996. Ecological correlates to social structure in two lemur species in Madagascar. American Journal of Physical Anthropology 100:487-506.
Overdorff, D. J., A. Merelender, P. Talata, A. Telo, and Z. Forward. In review. Demography and life history of Eulemur fulvus rufus in southeastern Madagascar from 1988 to1995.
Pulliam, H. R. 1988. Sources, sinks, and population regulation. American Naturalist 132:652-661.
Raymond, I. 1995. Approche phytoecologique sur l'evaluation qualitative et quantitative des utilisations villageoises des resources naturelles en forêt dense humide sempervierente. DEA Thesis. Universite d'Antananarivo, Antananarivo, Madagascar.
Richard, A. F., P. Rakotomanga, and M. Schwartz. 1991. Demography of Propithecus verreauxi at Beza Mahafaly, Madagascar: sex ratio, survival, and fertility, 1984-1988. American Journal of Physical Anthropology 84: 307-322.
Rigamonti, M. M. 1993. Home range and diet in red ruffed lemurs (Varecia variegata rubra) on the Masoala Peninsula, Madagascar. Pages 25-39 in P. M. Kappeler and J. U. Ganzhorn, editors. Lemur social systems and their ecological basis. Plenum Press, New York, New York, USA.
Smith, A. P., N. Horning, and D. Moore. 1997. Regional biodiversity planning and lemur conservation with GIS in western Madagascar. Conservation Biology 11: 498-512.
Southwick, C. H., and F. C. Cadigan. 1972. Population studies of Malaysian primates. Primates 13:1-18.
Stapanian, M. A., S. P. Cline, and D. L. Cassell. 1997. Evaluation of a measurement method for forest vegetation in a large-scale ecological survey. Environmental Monitoring and Assessment 45(3):237-257.
Sterling, E. J. 1993. Patterns of range use and social organization in aye-ayes (Daubentonia madagascariensis) on Nosy Mangabe. Pages 1-9 in P. M. Kappeler and J. U. Ganzhorn, editors. Lemur social systems and their ecological basis. Plenum Press, New York, New York, USA.
Sterling, E. J., and N. Rakotoarison. 1998. Rapid assessment of primate species richness and density on Masoala peninsula, eastern Madagascar. Folia Primatologica 69(suppl.1).
Struhsaker, T. T. 1975. The red colobus monkey. Chicago University Press, Chicago, Illinois, USA.
Sussman, R W. 1991. Demography and social organization of free-ranging Lemur catta in the Beza Mahafaly Reserve, Madagascar. American Journal of Physical Anthropology 84(1):43-58.
Tattersall, I. 1982. The primates of Madagascar. Columbia University Press, New York, New York, USA.
Taylor, B. L., and T. Gerrodette. 1993. The uses of statistical power in conservation biology: The Vaquita and Northern Spotted Owl. Conservation Biology 7(3):489-500.
Terborgh, J. 1983. Five new world primates. Princeton University Press, Princeton, New Jersey, USA.
Vasey, N. 1996. Feeding and ranging behavior of red ruffed lemurs (Varecia variegata rubra) and white fronted lemurs (Lemur fulvus albifrons). American Journal of Physical Anthropology Suppl. 22:234-235.
Walters, C. J., and C. S. Holling. 1990. Large-scale management experiments and learning by doing. Ecology 71(6):2060-2068.
Watson, S, J. Ward, K. Izard, and D. Stafford. 1996. An analysis of birth sex ratio bias in captive prosimian species. American Journal of Primatology 38(4): 303-314.
Wells, M., and K. E. Brandon. 1993. The principles and practice of buffer zones and local participation in biodiversity conservation. Ambio 22:157-162.
Wells, M., K. Brandon, and L. Hannah. 1992. People and parks: linking protected area management with local communities. The World Bank publications, Washington, D.C., USA.
White, F. J. 1989. Diet, ranging behavior, and social organization of the black and white ruffed lemur Varecia variegata variegata in Southeastern Madagascar. American Journal of Physical Anthropology 78(2):323.
White, F. J., D. J. Overdorff, E. A. Balko, and P. C. Wright. 1995. Distribution of ruffed lemurs (Varecia variegata) in Ranomafana National Park, Madagascar. Folia Primatologica 64(3):124-131.
Whitesides, G. E., J. F. Oates, S. M. Green, and R. P. Kluberdanz. 1988. Estimating primate densities from transects in a West African rain forest: a comparison of techniques. Journal of Animal Ecology 57(2): 345-368.
World Bank, United States Agency for International Development, Cooperation Suisse, UNESCO, United Nations Development Program, and Worldwide Fund for Nature. 1988. Madagascar environmental action plan. Volume 1. World Bank, Washington, D.C., USA.
Zielinski, W. J., and H. B. Stauffer. 1996. Monitoring Martes populations in California; survey design and power analysis. Ecological Applications 6(4):1254-1267.
Address of Correspondent:
Adina Merenlender
Environmental Science, Policy, and Management Department
University of California, Berkeley
Berkeley, CA, USA 94720-3110
Phone: (707) 744-1270
Fax: (707) 744-1040
adina@nature.berkeley.edu
*The copyright to this article passed from the Ecological Society of America to the Resilience Alliance on 1 January 2000.
![]()